Para
el estudio de la variabilidad genética
de la raza se obtuvieron muestras sanguíneas
de 16 animales (14 hembras y 2 machos), y
se procedió a la extracción
del ADN según protocolo estándar
con mezcla fenol-cloroformo (Ausubel et al.,
1987). Los 15 marcadores de ADN de tipo microsatélite
utilizados fueron: CSSM66, ETH10, ETH152,
ETH225, ETH3, HEL1, HEL5, HEL9, ILST5 INRA23,
INRA35 INRA37, INRA5, INRA63 y TGLA44, todos
ellos marcados fluorescentemente. Los productos
amplificados de PCR (Reacción en
Cadena de la Polimerasa) fueron analizados
mediante electroforesis capilar, con un secuenciador
de ADN Applied Biosystems 3100 (ABI
3100), e interpretados posteriormente mediante
el software de análisis GENESCAN, utilizando
para ello un marcador estándar de tamaños
con fluorescencia TAMRA 350.
El análisis genético-estadístico
se realizó a partir de las frecuencias
alélicas aportadas por los marcadores.
Se calcularon los estadísticos de variabilidad
genética, heterocigosidad esperada
(He) y observada (Ho) y se realizó
el test de equilibrio de Hardy-Weinberg usando
el método de la Cadena de Markov mediante
el programa GENEPOP (Raymond and Rousset,
1995). Así mismo, se calculó
el índice de contenido polimórfico
(PIC) y la probabilidad de exclusión
de cada uno de los marcadores y su combinada
(PE). La estructura genético-poblacional
se analizó mediante los F-estadísticos,
implementados en el programa FSTAT (Goudet,
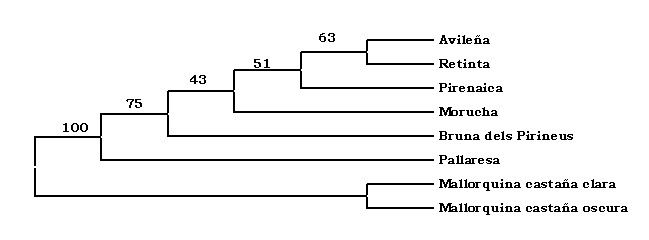
2000). Para el estudio de las relaciones genéticas
entre las razas y la posterior construcción
del dendrograma se utilizó la distancia
DA de Nei (Nei et al.,
1983) y el algoritmo NJ (neighbourn-joining
method) (Saitou and Nei, 1987), usando
para ello el programa DISPAN (Ota, 1993).
Como referencia y entronque para el diseño
de las relaciones filogenéticas de
la raza Pallaresa, se utilizó la información
de las frecuencias génicas de otras
razas españolas (Avileña,
Bruna dels Pirineus, Mallorquina, Morucha,
Pirenaica y Retinta) procedentes
de los trabajos realizados por Cañón
et al. (2001) y Aranguren-Méndez et
al. (2002).
Todos
los marcadores amplificaron correctamente
y fueron polimórficos en la raza Pallaresa.
El promedio de diversidad genética
poblacional (He), sobre todos los loci, fue
de 0,663±0,043, valor de variabilidad
genética comparable al obtenido en
otras razas bovinas europeas (Kantanen et
al., 2000; Cañón et al., 2001).
El índice de contenido polimórfico
(PIC) y la probabilidad de exclusión
de cada uno de los marcadores y su combinada
(PE) también fueron comparables a otras
poblaciones, siendo el valor de PE combinada
del 99,9%, confirmando los marcadores genéticos
utilizados, su utilidad, tanto para la identificación
individual como para la verificación
de paternidades.
El déficit global de heterocigotos
fue del 13,6% (P<0,001), aunque únicamente
4 loci (HEL1, INRA5, INRA35 e INRA37) contribuyeron
al mismo, por lo que no podemos atribuir a
la consanguinidad ser la causa principal que
haya ocasionado dicho déficit. La presencia
de alelos nulos no detectables podría
explicar parte del importante déficit
de heterocigotos observado en estos marcadores,
tal como ha sido reportado previamente en
otros trabajos (Høj Petersen and Bendixen,
2000; Kantanen et al., 2000).
Actualmente
todos los representantes de la raza bovina
Pallaresa están ubicados en un único
rebaño. Lógicamente, hubiéramos
esperado que el déficit de heterocigotos
observado y causado por la consanguinidad
(debido a un mayor grado de apareamiento entre
individuos emparentados) fuera realmente importante.
Sin embargo, y tal como podemos ver por los
resultados obtenidos esto no ha sido así.
Una posible explicación a que esta
población de reducido tamaño
censal se halle mayoritariamente en equilibrio
genético de Hardy-Weinberg, podría
deberse al hecho de que dicho rebaño
se ha ido constituyendo, en las últimas
décadas, a partir de la reagrupación
de individuos, coincidentes con el prototipo
racial (principalmente el color blanco uniforme
de su capa), de diferentes lugares y propietarios
de estos valles pirenaicos (comunicación
oral). La diferente procedencia de algunos
de los individuos (generalmente los más
viejos), y la más que probable aportación
genética, en algún determinado
momento, por parte de individuos de la raza
Bruna dels Pirineus (comparten rebaño
y pastos de puerto), habrían ayudado
a mantener los niveles de heterocigosidad
y variabilidad genética en esta reducida
y localizada población.
Y
ya por último, el dendrograma nos muestra,
de forma gráfica, la estrecha relación
existente entre la vaca Pallaresa y la Bruna
dels Pirineus, así como la relación
con otras razas peninsulares y la insular
Mallorquina. Los valores en los puntos de
bifurcación del árbol representan
los porcentajes de replicaciones bootstrap
después de 1.000 permutaciones.
Locus |
Alelos |
He |
Ho |
PIC |
PE |
FIS |
| CSSM66 |
8 |
0,85 |
0,75 |
0,80 |
0,75 |
-0,120*** |
| ETH10 |
5 |
0,74 |
0,75 |
0,67 |
0,48 |
-0,020*** |
| ETH152 |
5 |
0,71 |
0,69 |
0,64 |
0,44 |
-0,038*** |
| ETH225 |
5 |
0,72 |
0,62 |
0,66 |
0,47 |
-0,140*** |
| ETH3 |
5 |
0,78 |
0,75 |
0,74 |
0,50 |
-0,040*** |
| HEL1 |
4 |
0,72 |
0,37 |
0,64 |
0,44 |
-0,487*** |
| HEL5 |
7 |
0,68 |
0,81 |
0,62 |
0,44 |
-0,200*** |
| HEL9 |
4 |
0,65 |
0,69 |
0,57 |
0,37 |
-0,054*** |
| ILST5 |
2 |
0,17 |
0,19 |
0,15 |
0,08 |
-0,071*** |
| INRA5 |
4 |
0,65 |
0,44 |
0,57 |
0,37 |
-0,331*** |
| INRA23 |
5 |
0,80 |
0,69 |
0,74 |
0,56 |
-0,147*** |
| INRA35 |
4 |
0,46 |
0,12 |
0,40 |
0,24 |
-0,733*** |
| INRA37 |
6 |
0,55 |
0,25 |
0,50 |
0,33 |
-0,556*** |
| INRA63 |
4 |
0,77 |
0,87 |
0,69 |
0,50 |
-0,144*** |
| TGLA44 |
6 |
0,68 |
0,62 |
0,60 |
0,40 |
-0,083*** |
Media |
4,9±0,4 |
0,66±0,04 |
0,57±0,06 |
|
0,99 |
0,136±0,061 |

|