Per
a l'estudi de la variabilitat genètica
de la raça es varen obtenir mostres
sanguínies de 16 animals (14 femelles
i 2 mascles), i es procedí a l'extracció
de l'ADN segons protocol estàndard
amb mescla fenol-cloroform (Ausubel et al.,
1987). Els 15 marcadors d'ADN de tipus microsatèl.lit
utilitzats foren: CSSM66, ETH10, ETH152, ETH225,
ETH3, HEL1, HEL5, HEL9, ILST5 INRA23, INRA35
INRA37, INRA5, INRA63 i TGLA44, tots ells
marcats fluorescentment. Els productes amplificats
de la PCR (Reacció en Cadena de
la Polimerasa) foren analitzats mitjançant
electroforesi capil•lar, amb un seqüenciador
d'ADN Applied Biosystems 3100 (ABI
3100), i interpretats posteriorment amb el
software d'anàlisi GENESCAN, utilitzant
un marcador estàndard de tamanys amb
fluorescència TAMRA 350.
L'anàlisi
genètic-estadístic es realitzà
a partir de les freqüències gèniques
aportades pels marcadors. Es varen calcular
els estadístics de variabilitat genètica,
heterozigositat esperada (He) i observada
(Ho) i es realitzà el test d'equilibri
de Hardy-Weinberg utilitzant el mètode
de la Cadena de Markov mitjançant el
programa GENEPOP (Raymond and Rousset, 1995).
Així mateix, es calculà l'índex
de contingut polimòrfic (PIC) i la
probabilitat d'exclusió de cadascun
dels marcadors i la seva combinada (PE). L'estructura
genètica-poblacional s'analitzà
mitjançant els F-estadístics,
implementats en el programa FSTAT (Goudet,
2000). Per a l'estudi de les relacions genètiques
entre les races i la posterior construcció
del dendrograma es va utilitzar la distància
DA de Nei (Nei et al.,
1983) i l'algoritme NJ (neighbourn-joining
method) (Saitou and Nei, 1987), implementats
en el programa DISPAN (Ota, 1993). Com a referència
i entroncament per al disseny de les relacions
filogenètiques de la raça Pallaresa,
s'utilitzà la informació de
les freqüències gèniques
d'altres races espanyoles (Avileña,
Bruna dels Pirineus, Mallorquina, Morucha,
Pirenaica i Retinta) procedents
dels treballs realitzats per Cañón
et al. (2001) i per Aranguren-Méndez
et al. (2002).
Tots
els marcadors varen amplificar correctament
i foren polimòrfics en la raça
Pallaresa. La mitjana de diversitat genètica
poblacional (He), sobre tots els loci, fou
de 0,663±0,043, valor de variabilitat
genètica comparable a l'obtingut en
altres races bovines europees (Kantanen et
al., 2000; Cañón et al., 2001).
L'índex de contingut polimòrfic
(PIC) i la probabilitat d'exclusió
de cadascun dels marcadors i la seva combinada
(PE) també foren comparables a altres
poblacions, essent el valor de PE combinada
del 99,9%, confirmant els marcadors genètics
utilitzats, la seva utilitat, tant per a la
identificació individual com per a
la verificació de paternitats.
El
dèficit global d'heterozigots fou del
13,6% (P<0,001), encara que únicament
4 loci (HEL1, INRA5, INRA35 i INRA37) contribuïren
al mateix, pel que no podem atribuir a la
consanguinitat ser la causa principal que
hagi ocasionat aquest dèficit. La presència
d'al.les nuls no detectables podria explicar
part de l'important dèficit d'heterozigots
observat en aquests marcadors, tal com ja
ha sigut reportat prèviament en altres
treballs (Høj Petersen and Bendixen,
2000; Kantanen et al., 2000).
Actualment,
tots els representants de la raça bovina
Pallaresa estan ubicats en un únic
ramat. Lògicament, hauríem esperat
que el dèficit d'heterozigots observat
i causat per la consanguinitat (degut a un
major grau d'aparellament entre individus
emparentats) fos realment important. No obstant,
i tal com podem veure pels resultats obtinguts
això no ha sigut així. Una possible
explicació a que aquesta població
de reduït tamany censal es trobi majoritàriament
en equilibri genètic de Hardy-Weinberg,
podria deures al fet de que aquest ramat s'ha
anat constituint, en les darreres dècades,
a partir de la reagrupació d'individus,
coincidents amb el prototip racial (principalment
el color blanc uniforme del seu pelatge),
de diferents llocs i propietaris d'aquestes
valls pirinenques (comunicació oral).
La diferent procedència d'alguns individus
(generalment els més vells), i la més
que probable aportació genètica,
en algun moment puntual, per part d'individus
de la raça Bruna dels Pirineus (comparteixen
ramat i pastures de port), haurien ajudat
a mantenir els nivells d'heterozigositat i
variabilitat genètica en aquesta reduïda
i localitzada població.
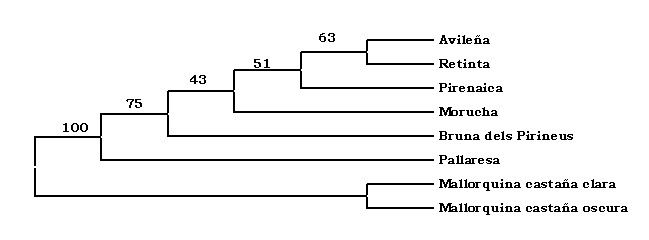
I
ja per últim, el dendrograma ens mostra,
de forma gràfica, l'estreta relació
existent entre la vaca Pallaresa i la vaca
Bruna dels Pirineus, així com la relació
amb altres races peninsulars i l'insular Mallorquina.
Els valors en els punts de bifurcació
de l'arbre representen els percentatges de
replicacions bootstrap desprès de 1.000
permutacions.
Locus |
Al.lels |
He |
Ho |
PIC |
PE |
FIS |
| CSSM66 |
8 |
0,85 |
0,75 |
0,80 |
0,75 |
-0,120*** |
| ETH10 |
5 |
0,74 |
0,75 |
0,67 |
0,48 |
-0,020*** |
| ETH152 |
5 |
0,71 |
0,69 |
0,64 |
0,44 |
-0,038*** |
| ETH225 |
5 |
0,72 |
0,62 |
0,66 |
0,47 |
-0,140*** |
| ETH3 |
5 |
0,78 |
0,75 |
0,74 |
0,50 |
-0,040*** |
| HEL1 |
4 |
0,72 |
0,37 |
0,64 |
0,44 |
-0,487*** |
| HEL5 |
7 |
0,68 |
0,81 |
0,62 |
0,44 |
-0,200*** |
| HEL9 |
4 |
0,65 |
0,69 |
0,57 |
0,37 |
-0,054*** |
| ILST5 |
2 |
0,17 |
0,19 |
0,15 |
0,08 |
-0,071*** |
| INRA5 |
4 |
0,65 |
0,44 |
0,57 |
0,37 |
-0,331*** |
| INRA23 |
5 |
0,80 |
0,69 |
0,74 |
0,56 |
-0,147*** |
| INRA35 |
4 |
0,46 |
0,12 |
0,40 |
0,24 |
-0,733*** |
| INRA37 |
6 |
0,55 |
0,25 |
0,50 |
0,33 |
-0,556*** |
| INRA63 |
4 |
0,77 |
0,87 |
0,69 |
0,50 |
-0,144*** |
| TGLA44 |
6 |
0,68 |
0,62 |
0,60 |
0,40 |
-0,083*** |
Mitjana |
4,9±0,4 |
0,66±0,04 |
0,57±0,06 |
|
0,99 |
0,136±0,061 |

|